Quand on parle de consanguinité dans l’élevage du chien de race, le débat se transforme souvent en guerre de clans : les “pour” (fixer le type) et les “contre” (risques pour la santé). C’est une impasse. La consanguinité n’est ni morale ni immorale : c’est un levier génétique. Et comme tout levier, elle a un coût. Le vrai sujet est donc simple : quel niveau de risque accepte t-on, et surtout pour quoi faire.
Cet article met les choses à plat : il explique ce qu’est réellement la consanguinité, ce qu’elle mesure (et ne mesure pas), pourquoi le taux de consanguinité “5 générations” sous-estime souvent la réalité, et quelles pratiques limitent le risque sans tomber dans les dogmes.
Cet article consacré à la consanguinité du chien de race est un des plus complets que vous pourrez trouver sur internet, en français. Couvrir tous les volets de la consanguinité dans un seul article entraîne immanquablement une certaine longueur… C’est volontairement que je n’ai pas souhaité éclater cet article en plusieurs plus petits articles car il y a une progression logique dans l’apprentissage de ce sujet, qui n’aurait pas pu être assurée autrement. Par conséquent, n’hésitez pas à revenir et utiliser le sommaire qui vous permettra d’accéder directement aux différents chapitres au fur et à mesure de votre progression.
Sommaire :
- Préambule
- Introduction à la génétique
- Le taux de consanguinité
- Comment calculer un taux de consanguinité
- Que dit le taux de consanguinité ?
- Ligne directe et ligne colatérale
- Les effets de la consanguinité
- Combien de générations dois-je prendre en compte dans le calcul du coefficient de consanguinité ?
- LOF Select
- Les différentes options de calcul du taux de consanguinité
- Sous-estimation des taux de consanguinité issus des pédigree
- SNP (Single Nucleotide Polymorphism)
- RoH – Segments d’homozygotie
- La recombinaison génétique
- Le déséquilibre de liaison
- Consensus sur les méthodes de calcul ROH
- Embark
- Le coefficient AVK
- Les PedCharts
- Le coefficient de parenté (Kinship)
- Les véritables niveaux de consanguinité des chiens de race
- Les niveaux de consanguinité chez les autres animaux domestiques
- L’hypertype
- La consanguinité du chien de race : la roulette Russe ?
- Quelle est la valeur de consanguinité à ne pas dépasser pour le chien de race ?
- Dois-je quand même me préoccuper du coefficient de consanguinité si je fais les tests de santé disponibles pour ma race ?
- Conduite d’élevage à adopter
- La mauvaise utilisation de la consanguinité dans les pratiques d’élevage
- Les traits polygéniques
- Les reproducteurs populaires
- Conclusion
Préambule
Le bon vieux temps
La plupart du temps, nous élevons toujours les chiens de la même manière qu’il y a 100 ans. Nous examinons les pédigrees, nous apprenons à connaître les chiens à partir de lignées particulières, nous examinons la qualité de la progéniture que produisent les chiens et nous évaluons les bons et les mauvais points des mâles et des femelles reproducteurs.
Bien que le processus général de reproduction reste le même, le contexte a radicalement changé au fil des décennies. Il y a cent ans, les races étaient fraîchement créées à partir d’un stock qui a été élevé en grande partie pour la fonction plutôt que pour l’apparence. Avant l’avènement du livre des origines, vous pouviez introduire un peu d’autres races chaque fois que vous pensiez qu’elle pourrait compléter les caractéristiques que vous recherchiez. Lorsque les clubs de races ont été créés, nous avons commencé à restreindre l’élevage aux chiens enregistrés seulement afin que l’ascendance soit connue et pour augmenter la valeur des chiens de « race pure ». Cela a conduit à fermer le livre des origines. La plupart des éleveurs n’y pensaient probablement pas beaucoup à l’époque, et nos connaissances de la génétique et de l’hérédité n’étaient que rudimentaires, mais la généalogie fermée avait des conséquences importantes parce qu’elle empêchait l’introduction de gènes frais dans la race. En fait, tous les gènes qu’une race aurait jamais eu se trouvent dans les premiers chiens, ceux de la population fondatrice de la race. Les gènes des chiens de race pure modernes sont des copies de gènes qui étaient présents chez les chiens fondateurs de la race, et ils ont été dupliqués et transmis aux descendants, génération après génération.
Le problème avec un pool de gènes fermé est que les gènes peuvent être perdus ou supprimés, mais aucun nouveau gène ne peut être ajouté. Au fil des générations, les gènes pour les traits indésirables sont éliminés par sélection, mais d’autres gènes sont perdus juste par hasard (dérive génétique) ou comme conséquence involontaire de la sélection des traits. Tôt ou tard, des gènes qui sont critiques pour certaines fonctions vitales sont perdus, et lorsque cela se produit, il peut y avoir des conséquences négatives. Les maladies génétiques, la faible fertilité, les problèmes de comportement et d’autres problèmes commencent à apparaître dans la race, et l’élevage de chiens en bonne santé devient de plus en plus difficile. Voilà où nous en sommes aujourd’hui avec de très nombreuses races.
L’élevage du chien de race est basé sur la consanguinité.
Au début de la création d’une race, on accouple deux chiens de races différentes. Ces deux sujets sont supposés être, en théorie, non apparentés. La première génération produit une portée avec un taux de consanguinité de 0% car ils n’ont (toujours en théorie) aucun ancêtre commun.
Par la suite, des combinaisons hautement consanguines (père-fille / mère-fils / frère et soeur) sont effectuées afin de fixer le type attendu de la race (traits physiques et caractère). D’autres chiens peuvent venir de l’extérieur (toujours considérés comme étant non apparentés en théorie) et s’accoupler avec le fruit des premières générations. Une fois que les traits physiques et le caractère correspondent aux attentes, le standard de la race est publié et le livre des origines est fermé.
Les éleveurs de chiens de race ne peuvent par la suite que reproduire en consanguinité.
Introduction à la génétique
Toutes les cellules de l’organisme contiennent des chromosomes qui sont formées par de l’ADN compacté. Les chromosomes se situent dans le noyau des cellules.
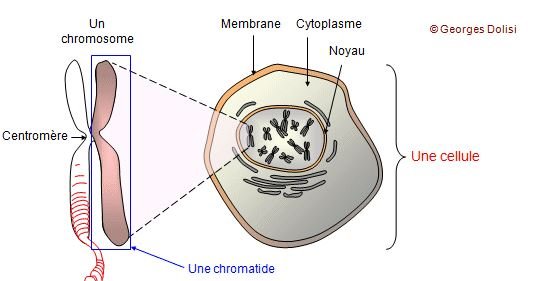
Le chromosome est le support de l’information génétique ou de l’hérédité.
Le chien possède 39 paires de chromosomes (38 paires d’autosomes et 1 paire de chromosomes sexuels). Chaque paire de chromosome est constituée d’un chromosome d’origine paternelle et d’un chromosome d’origine maternelle.
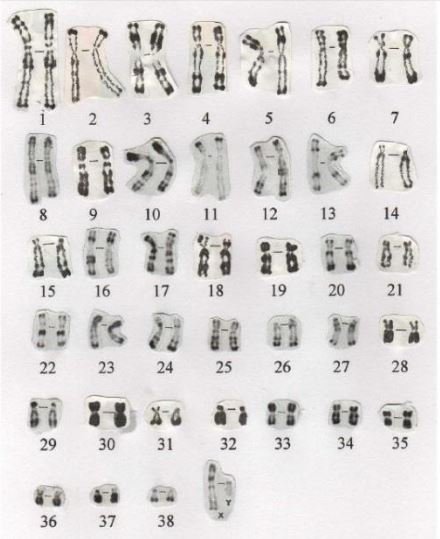
L’homme possède 23 paires de chromosomes. Il ne semble pas y avoir d’explications au nombre de chromosomes trouvés dans une espèce. Le kangourou en possède 16 tout comme le parasite schistosome, l’escargot et la tomate en ont tous les deux 24, la girafe et la pistache en ont 30, le porc-épic et l’artichaut en ont 34, la souris et la mangue en ont 40, le rat et l’avoine en ont 42, le hérisson et la fougère à raisin « Sceptridium » en ont 90, le papillon bleu atlas en a environ 500 tandis que le cerf aboyeur n’en a que 6 (chez les femelles) ou 7 (chez les mâles)…
Chacun des 39 chromosomes est en réalité de l’ADN condensé, qui se compacte avec des protéines.
L’ADN est subdivisé en petite portions que l’on appelle des gènes. Un gène est une position sur la molécule d’ADN et il existe différentes versions de gène que l’on appelle des allèles. Un gène porte l’information à l’origine d’un caractère héréditaire (couleur des yeux, des poils etc). Chaque gène se présente sur les deux chromosomes (celui hérité du père et celui hérité de la mère) au même locus (même endroit) sur la paire de chromosomes. Un allèle est une version différente d’un même gène. Les allèles sont à l’origine de la diversité génétique d’un chien et elles définissent son génotype.
Génotypes et phénotypes
Le génotype est le mélange particulier d’allèles qu’un individu présente sur ses chromosomes. Chez les chiens, il y a un total d’environ 20.000 gènes répartis sur les 39 paires de chromosomes. Pour chaque gène, un individu hérite d’une copie de la mère (sur le chromosome issue de la mère) et d’une copie du père (sur le chromosome issue du père). L’emplacement sur le chromosome où se trouve le gène est appelé locus (loci au pluriel).
Le phénotype d’un animal est ce que vous considéreriez comme un trait (une caractéristique) – taille, couleur, tempérament, vitesse de course, fréquence cardiaque, fertilité, etc. La plupart des traits d’un animal sont polygéniques (déterminés par plusieurs gènes) et sont la conséquence des effets à la fois des gènes et de l’environnement sur un animal. Par exemple, la taille d’un chien à l’âge adulte sera déterminée par les gènes qu’il a hérité de ses parents, mais l’alimentation qu’il a eu lorsqu’il était chiot aura également une incidence sur sa taille finale. Le trait « taille » est donc une conséquence des gènes particuliers hérités des parents et des facteurs non génétiques – dans ce cas, l’alimentation pendant la période de croissance.
Différence entre gènes et allèles?
Un gène possède différentes versions appelées allèles.
Pour les traits ou les maladies monogéniques (impliquant un seul gène), c’est la variation allélique du gène impliqué qui déterminera le trait ou la maladie.
Pour les traits ou les maladies polygéniques (impliquant plusieurs gènes), c’est la combinaisons des variations alléliques de l’ensemble des gènes impliqués qui déterminera le trait ou la maladie.
Allèles homozygotes et allèles hétérozygotes
Si les allèles hérités de chaque parent sont exactement les mêmes, le chien est homozygote pour ce gène. Si les allèles sont différents, le chien est hétérozygote pour ce même gène.
Allèles dominants et allèles récessifs
Chaque organisme possède au minimum deux allèles pour chaque gène.
Dans un scénario simple, la combinaison des allèles hérités de chaque parent ont une relation de dominance ou de récessivité dans l’expression d’une mutation.
Ainsi, la variation allélique dominante héritée d’un seul des deux parents est suffisante pour que la mutation du gène s’exprime.
Alors que la variation allélique récessive nécessite d’être héritée des deux parents pour que la mutation du gène puisse s’exprimer.
Les bases de la génétique
Allez, une dernière petite vidéo, en Français qui résume les points importants des bases de la génétique.
Le taux de consanguinité
“Le taux de consanguinité est la probabilité que les deux allèles d’un gène à un locus pris au hasard, soient identiques (homozygotes) par descendance.”
Il permet également de prédire l’homozygotie génétique d’un chien.
Il existe plusieurs formules de calculs de coefficient de consanguinité.
La plus ancienne (1922) est Wright puis en 1948 Malecot, en 1962 Kudo, en 1997 Ballou puis récemment en 2000, Hardiman (limité à 5 générations).

Nous allons ici développer le coefficient de consanguinité inventé par Sewall Wright .
Il était un généticien de l’évolution et a mis au point une formule qui est à ce jour la plus répandue et la plus utilisée
Le pedigree du chien, mais plus globalement, le livre fermé des origines de chaque race de chiens est la principale source d’information pour pouvoir appliquer la formule de Wright. On parle donc de coefficient de consanguinité généalogique et non de coefficient de consanguinité génétique (que nous développerons plus loin).
Comment calculer un taux de consanguinité
La vidéo suivante vous apprendra à calculer un taux de consanguinité depuis un pédigree 3 générations en utilisant la méthode dite « des chemins ».
Que dit le taux de consanguinité ?

Vous êtes dorénavant incollable sur le calcul du taux de consanguinité (COI – Coefficient of Inbreeding en anglais).
Mais gardez à l’esprit que :
- Le taux de consanguinité ne donne pas d’information sur l’identité des gènes qui seront homozygotes. En effet, ils peuvent être de bons gènes comme de mauvais gènes…
- Il ne dit pas si les effets de la consanguinité seront bons ou mauvais.
- Il ne prédit pas si les chiots seront en bonne ou en mauvaise santé.
- Le taux de consanguinité est une estimation de la consanguinité moyenne attendue pour une portée. Mais la consanguinité individuelle pour chaque chiot au sein de la portée peut être supérieure ou inférieure à cette moyenne.
- Le taux de consanguinité est une probabilité statistique de l’homozygotie attendue d’un individu. Si l’homozygotie apparaît sur un gène désirable, alors le taux de consanguinité aura été une mesure de bénéfice potentiel. Si l’homozygotie apparaît sur un gène indésirable, alors le taux de consanguinité aura été une mesure de risque potentiel.
La consanguinité augmente la probabilité que, sur un locus donné, un chien ait deux copies identiques d’un allèle héritées d’un ancêtre commun. Dit autrement : elle augmente l’homozygotie par descendance. Cette homozygotie peut “figer” des traits visibles… mais elle peut aussi révéler des combinaisons délétères (et pas seulement des maladies monogéniques faciles à dépister).
Si on élève dans une population dont le livre des origine est fermé, tu ne peux pas échapper à une réalité : la diversité génétique se perd. Tu peux seulement choisir à quelle vitesse.
Ligne directe et ligne colatérale
La consanguinité en ligne directe ou In breeding est l’accouplement de deux individus ayant entre eux, un lien de parenté étroit. Par exemple l’accouplement père – fille, frère – sœur, oncle et nièce etc. Ces mariages sont désormais interdits par la SCC, sauf dérogations.
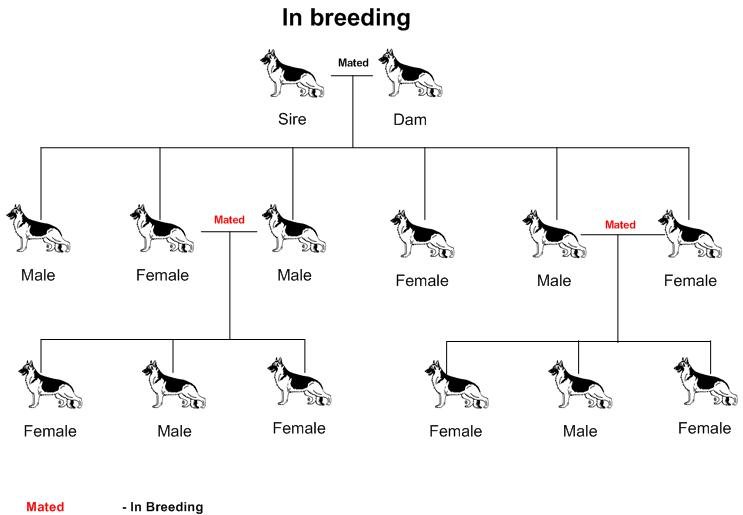
La consanguinité en ligne collatérale, également appelée Line breeding est l’accouplement de deux individus ayant au moins un ancêtre commun.
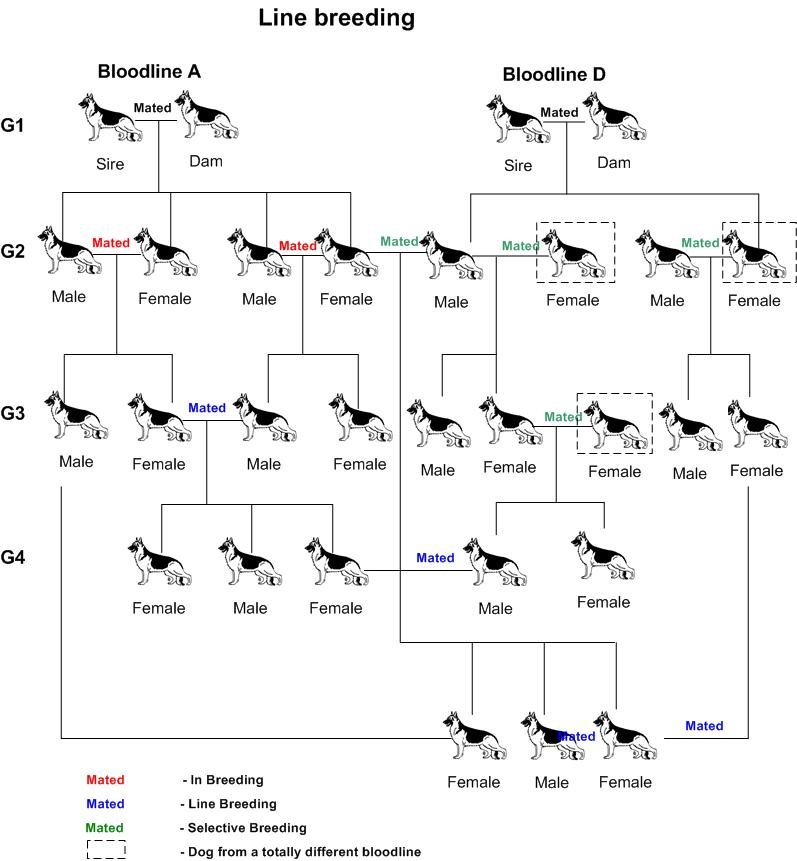
Quel que soit leurs noms, il s’agit dans tous les cas de combinaisons consanguines à différents degrés. Par conséquent elles alimentent la dépression de consanguinité chez les chiens de race.
Les effets de la consanguinité
Lorsque les éleveurs utilisent la consanguinité pour leur programme de reproduction, ils risquent également une longue liste de dommages applicables aux animaux. Aussi bien sur le court terme que sur le long terme. En conséquence l’uniformité et la transmission de caractéristiques spécifiques d’un individu à un autre augmentent avec le temps. Cependant, il en va de même de l’infertilité, de la mortalité des descendants, de l’espérance de vie, des maladies génétiques héréditaires et autres maladies dérivées.
Quand le livre généalogique est fermé, des gènes du patrimoine génétique de la race sont perdus par sélection et par «dérive génétique» (due au hasard).
Si vous démarrez une race avec un pool de gènes fondateurs et que vous fermez le livre des origines pour ne reproduire par la suite qu’en consanguinité, vous perdez des gènes à chaque génération. Ainsi les gènes ne pouvant être remplacés, le pool génétique s’épuise. Par conséquent, les animaux (quels qu’ils soient) qui évoluent dans des pools de gènes fermés finissent par disparaître.
| Avantages de la consanguinité | Inconvénients de la consanguinité |
|---|---|
| Plus d’uniformité dans la portée | Baisse de la fertilité |
| Plus d’aptitude à transmettre (reproducteurs dits « traceurs ») | Baisse de la vitalité |
| Fixation des traits physiques et comportementaux | Défauts de naissance |
| Réduction de la taille des chiots | |
| Portées moins nombreuses | |
| Croissance plus lente | |
| Mortalité néonatale plus élevée | |
| Baisse de longévité | |
| Augmentation du nombre de maladies génétiques | |
| Diminution de la diversité génétique |
Combien de générations dois-je prendre en compte dans le calcul du coefficient de consanguinité ?

Aucun ancêtre commun n’apparaît sur le pedigree cinq générations de votre chien ? Par conséquent vous pensez que son taux de consanguinité est de 0%, puisque d’ailleurs, c’est ce qui est écrit sur son pédigree ? Grave erreur…
TOUS les chiens de races pures sont consanguins.
Par définition une race de chien est considérée comme une population fermée et consignée dans des livres des origines. La plupart des races de chiens ont été créées avec une poignée d’individus en théorie non apparentés (les fondateurs) puis le livre des origines a été fermé. Par la suite les éleveurs ne peuvent reproduire qu’en consanguinité.
La consanguinité du chien de race augmente sans relâche car elle se cumule avec les générations.
Il est donc important de comprendre qu’une race possède une consanguinité significative très tôt dans le pedigree. Aussi, utiliser des pedigrees peu profonds (trois ou cinq générations) pour calculer un taux de consanguinité sous-estime très largement l’état homozygote d’un chien de race.
La lecture des pédigrees doit pouvoir se faire le plus profondément possible (jusqu’aux fondateurs).
Vous n’êtes pas convaincus ? Alors le meilleur moyen de vous convaincre est de commencer au commencement du pedigree, avec les premiers fondateurs de la race. Puis de calculer la consanguinité au travers des chemins pour les chiens de 8ème ou 10ème génération après le premier couple fondateur.
Si le coefficient de consanguinité dans la race était de 20% à la dixième génération après la création de la race, votre taux de consanguinité de 0%, une dizaine de générations plus tard est une idée fausse…
Gardez cela à l’esprit lorsque vous vous lancez dans le calcul de consanguinité des générations actuelles. D’autant plus lorsque vous consultez le taux de consanguinité de 5 générations proposé par la SCC sur son site LOF Select.
Un taux de consanguinité de 0% n’existe pas chez le chien de race…
Imaginons un iceberg : le taux de consanguinité 5G ne représente que la partie émergée de l’iceberg ! Le taux de consanguinité cinq générations sous-estime complètement l’état d’homozygotie de l’individu contrairement à un taux de consanguinité toutes générations, c’est à dire jusqu’aux fondateurs (ou en tous cas, le plus loin possible dans le pedigree).
La consanguinité ne se dilue pas avec le temps…
Au contraire, elle se cumule…

Pour estimer la hauteur totale de cet iceberg, vous devez prendre l’intégralité des données ; la partie émergée ET la partie immergée.
Le taux de consanguinité « toutes générations » est le taux le plus représentatif du niveau d’homozygotie d’un individu.
Avec uniquement un COI “5 générations”, on a une mesure courte, parfois utile, souvent trompeuse. Le danger : prendre un chiffre bas comme une preuve de diversité. Dans une race fermée, c’est souvent juste une preuve d’angle mort.
Le COI 5G n’est pas “faux”, il est incomplet. Si on le traite comme une vérité absolue, on pilote la race vers la perte de diversité sans s’en rendre compte.
LOF Select

Le pédigree de 5 générations est le pédigree le plus courant et le plus utilisé aujourd’hui. Il est délivré par les instances cynophiles (en France, la SCC – Société Centrale Canine) et accompagne toute vente de chiots LOF.
Un pédigree 5 générations implique 62 chiens.
Le pédigree 5G est utilisé par la majorité des éleveurs pour analyser les ancêtres d’un étalon lors de la préparation d’une future combinaison. Mais pourquoi 5 générations et pas plus ?
Parce qu’au delà de 5 générations, il est considéré par les instances cynophiles que l’influence d’un ancêtre situé au-delà de 5 générations est négligeable pour pouvoir influer sur le phénotype et le caractère. C’est-à-dire que la probabilité qu’un allèle se soit transmis depuis l’ancêtre commun se situant au-delà de la 5ème génération jusqu’au sujet considéré, par son côté paternel et maternel, est proche de zéro. Comprenez par là, qu’il y a peu de chances que les allèles du glorieux multi-champion de concours de beauté en 6ème génération puissent être hérités par votre chiot.
Cette profondeur de générations a également été choisie afin de pouvoir comparer les valeurs de consanguinité entre les sujets. En effet, la comparaison n’a de sens que si le calcul de consanguinité est effectué sur le même nombre de générations.
Cependant, le pédigree 5G ne reflète pas le statut homozygote de votre chien ! Les instances cynophiles sont presques exclusivement concentrées autour des questions du phénotype (les traits observables) et du cararactère (normal, puisque c’est ce qui définit la race d’un chien), mais sur un plan génétique, un pédigree si peu profond n’apporte que des informations erronées sur l’état homozygote des gènes de votre chien et donc sur les risques qui peuvent impacter sa santé…
La question de la santé est souvent reléguée en fin de réflexion. Lorsque les problèmes de santé se présentent, il est trop tard pour faire machine arrière. Alors, on cherche des tests de dépistage pour les nouvelles mutations récessives que les éleveurs ont eux-mêmes créé par consanguinité. Ainsi les éleveurs continuent d’alimenter eux-mêmes le vortex des maladies héréditaires.
Plus de 500 maladies héréditaires sont référencées à ce jour par OMIA
Les différentes options de calcul du taux de consanguinité

Il existe plusieurs options de calcul pour déterminer un coefficient de consanguinité à l’aide d’un logiciel dédié : “strictes” et “détendues”.
Stricte signifie que le logiciel doit strictement respecter la limite de génération d’ancêtres imposée pour le calcul du taux de consanguinité (par exemple calcul du taux de consanguinité sur les cinq dernières générations). Le logiciel n’utilisera pas les données au-delà de la limite définie même si les informations concernant les ancêtres sont disponibles.
Détendue signifie que le logiciel s’autorise à aller au-delà de la limite de générations fixée. Imaginez un pédigree de cinq générations dans lequel aucun ancêtre commun n’apparaît. Avec une option stricte, le taux de consanguinité serait de 0%@5G.
Cependant, si le logiciel est autorisé à regarder au-delà de la 5ème génération, il s’apercevrait par exemple, que le père d’un des ancêtres en 5ème génération apparait en 6ème génération des deux côtés du pédigree (côté père et côté mère). Cela permet au logiciel d’inclure une consanguinité supplémentaire due au père de cet ancêtre qu’il pourrait autrement ignorer.
Le calcul détendu génère des taux de consanguinité plus importants qu’avec les calculs strictes mais reflètent bien mieux la réalité…
Chaque option de calcul possède deux modes de calculs distinctifs :
- “Stricte avec consanguinité minimale d’ancêtre commun”
- “Stricte avec consanguinité maximale d’ancêtre commun”
- « Détendue avec consanguinité maximale”
- « Détendue avec vitesse maximale”
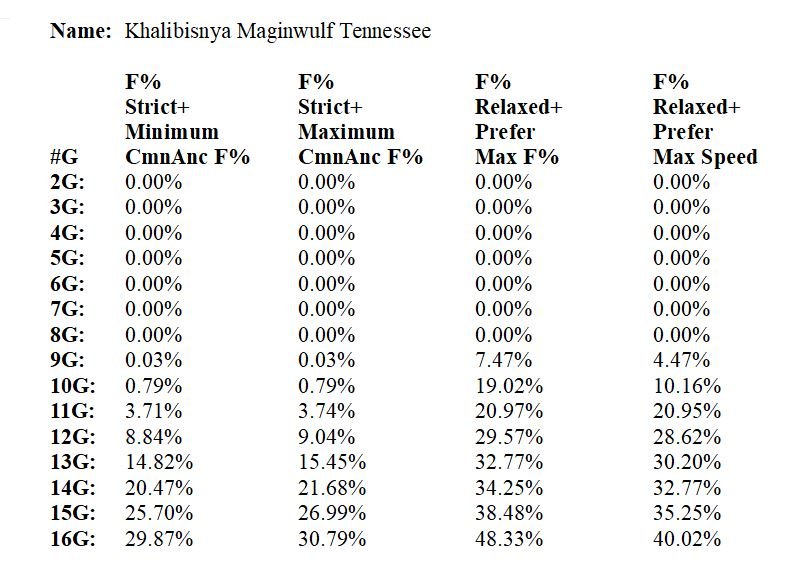
Ci-dessus, un exemple de calcul de taux de consanguinité effectué par un logiciel pour Khalibisnya Maginwulf Tennessee sur différentes profondeurs de pédigree (de 2 générations jusqu’à 16 générations) selon les quatres modes de calculs disponibles. Le mode de calcul « détendu avec consanguinité maximale » (3ème colonne) est à privilégier.
Il est maintenant évident que lorsque l’on compare des taux de consanguinité sur différents sites internet, leurs options de calcul doivent être identiques pour une comparaison pertinente.
Dans quel cas les valeurs de consanguinité seraient différentes d’un site à un autre ou d’un logiciel à un autre ?
- L’option de calcul choisie peut être différente
- La database utilisée par le logiciel de calcul de consanguinité peut contenir des pedigrees incomplets et/ou des données erronées.
Pour en savoir plus sur les différentes options de calcul des taux de consanguinité, le bouton de téléchargement ci-dessous vous permettra de comprendre le raisonnement appliqué pour chaque option au travers d’un exemple simple de pedigree.
Sous-estimation des taux de consanguinité issus des pédigree
Y a t-il une différence entre un taux de consanguinité généalogique (calculé depuis le pédigree) et un taux de consanguinité génétique (calculé depuis l’analyse ADN) ?
Les estimations généalogiques de la consanguinité en utilisant toutes les données du pedigree (jusqu’aux fondateurs de la race) sous-estiment généralement la consanguinité génétique. Parfois de beaucoup comme le démontre ce tableau ci-dessous. A gauche, les valeurs de consanguinité génétique, à droite les valeurs de consanguinité généalogique. Les valeurs sont à multiplier par 100 pour obtenir un pourcentage.
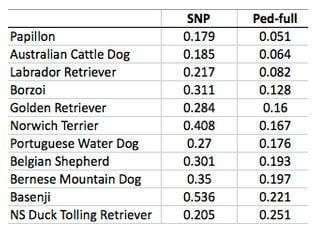
Les taux de consanguinité calculé depuis les pédigrées sont souvent sous-estimés en raison de toutes sortes d’erreurs.
– Les fondateurs d’une race sont toujours considérés comme étant non apparentés car de races différentes. Malheureusement, les races de chiens sont bien plus apparentées qu’on ne pourrait le penser sur le plan de l’ADN…
– Des erreurs « administratives » telles que des dates de naissance erronées, des ancêtres inconnus, des déclarations de saillies erronées, des périodes difficiles de l’histoire entraînant des informations manquantes – guerre 1939-1945 etc…
Tous ces manquements engendrent des liens incorrects dans le livre des origines et impactent le taux de consanguinité généalogique.
Néanmoins, les informations généalogiques ont été et continueront d’être utilisées pour estimer les paramètres génétiques des populations de chiens de race.
Les raisons sont les suivantes:
- Un vaste ensemble de marqueurs moléculaires (SNP > 200 000) au niveau du génome n’est devenu accessible que très récemment.
- Le coût du génotypage complet est trop élevé pour être supporté par tous les éleveurs et pour tous les chiens.
SNP (Single Nucleotide Polymorphism)
Les progrès de la science permettent dorénavant d’analyser les données ADN d’un chien. Cette nouvelle approche scientifique est une étape décisive dans la compréhension de la consanguinité. Pour connaître l’identité et la localisation des gènes sur les 39 paires de chromosomes du chien sans avoir à séquencer l’intégralité du génome (couteux et chronophage), les scientifiques utilisent ce qu’on appelle des marqueurs ADN.
Il existe principalement deux types de marqueurs :
- Les microsatellites (STR – Short Tandem Repeat connus également sous d’autres noms différents tels que SSLP – Simple Sequence Lenght Polymorphism, VNTR – Variable Number Tandem Repeat et SRR – Short Tandem Repeat)
- Les SNP (prononcez « snips » pour Single Nucleotide Polymorphism). Plus précis que les microsatellites, les SNP permettent d’identifier chacune des paires de bases (c’est à dire un seul nucléotide) à un endroit spécifique sur les chromosomes.
Les 39 paires de chromosomes du chien constitue son génome.
Le génome du chien est composé de milliards de paires de bases.
En temps normal, lorsque le génome est copié afin de créer une nouvelle cellule, il s’agit d’une copie parfaite. Mais ce n’est pas toujours le cas. En effet, une paire de bases peut être soustraite, rajoutée ou remplacée au lieu d’être copiée.
La substitution d’une paire de bases est appelée SNPs.
Il y a des millions de SNPs dans le génome du chien. Ce qui permet de différencier chaque chien. Certains SNP sont responsables des traits physiques. D’autres SNP sont impliqués dans les maladies. Comme les variations des paires de bases sont transmises de générations en générations, le ratio des différences entre l’ADN d’un chien et celui d’un autre chien peut définir leur degré d’apparenté.
RoH – Segments d’homozygotie
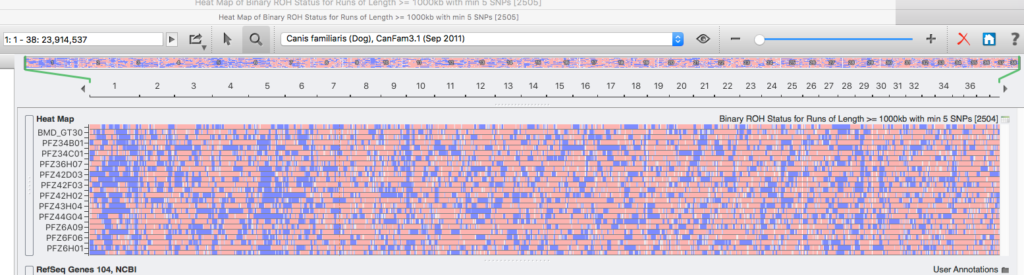
La méthode la plus courante pour déterminer la consanguinité à partir de l’analyse des SNP utilise une technique qui quantifie la consanguinité représentée dans les «segments d’homozygotie» (ROH – Runs of Homozygosity en anglais). Les ROH sont des régions des chromosomes où il existe de nombreux gènes homozygotes consécutifs.
Les gènes homozygotes produits par la consanguinité ne sont pas dispersés au hasard sur tous les chromosomes. Ils forment des blocs de gènes homozygotes dans les régions du chromosome où se trouvent des gènes sous sélection.
Si les gènes restent sous sélection continue au fil des générations, comme ce serait le cas pour les gènes de type dans une population de chiens de race pure, les blocs d’homozygotie auront tendance à s’allonger de plus en plus à chaque génération.
Parce que nous savons que ces blocs d’homozygotie se forment à la suite de la consanguinité, nous pouvons les utiliser pour estimer la fraction de consanguinité à travers le génome en additionnant la longueur totale des segments chez chaque chien et en divisant par la longueur totale des chromosomes.
La formule du calcul de la consanguinité estimé par ROH est égale à la somme des longueurs de segments d’homozygotie
divisé par la longueur totale des chromosomes
La recombinaison génétique
La consanguinité a tendance à produire des blocs d’homozygotie de plus en plus grands au fil du temps. Mais à chaque génération, il y a une étape dans la formation des gamètes (ovules et spermatozoïdes) appelée méiose, au cours de laquelle l’ADN est réarrangé par un événement appelé « recombinaison génétique ».
Pendant la recombinaison, des sections d’ADN se détachent et s’échangent entre la paire de chromosomes. Si ces cassures se produisent dans des segments d’homozygotie, les blocs seront divisés en morceaux plus petits. Par conséquent, nous nous attendrions à ce que les segments d’homozygotie deviennent plus court au fil des générations. Ainsi, nous pouvons établir que la consanguinité ancienne est représentée par des blocs de segments d’homozygotie courts alors que les blocs ROH les plus longs représentent la consanguinité récente.
Le pourcentage d’allèles transmis par les parents à sa progéniture ne se borne pas à une simple distribution égalitaire 50/50 .
En théorie, frères et soeurs sont génétiquement similaires à 25%, mais en réalité, ces valeurs ne sont que des moyennes. Il existe une variation significative entre le taux de consanguinité génétique du chien et le taux de consanguinité estimé à partir des pédigrees. En effet, en raison de la recombinaison, des différences d’hérédité peuvent se cumuler au fur et à mesure des générations. Ainsi, les résultats des taux de consanguinité basés sur la génétique peuvent varier jusqu’à 70 % par rapport aux taux de consanguinité basés sur les pédigrees !
L’analyse SNP est sans aucune mesure beaucoup plus efficace mais surtout beaucoup plus fiable que tous les autres coefficients calculés à partir des pédigrees.
Le déséquilibre de liaison
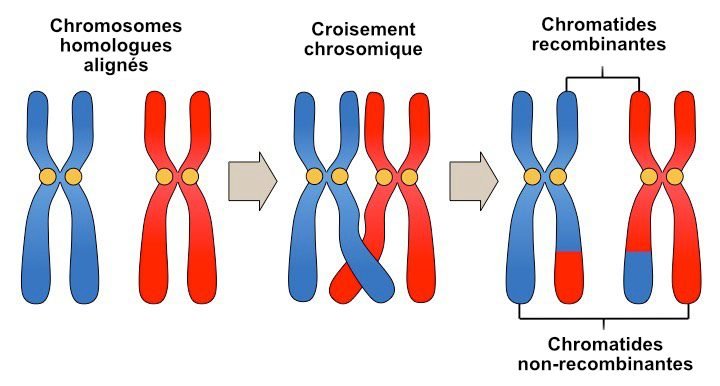
Outre l’utilité en tant que mesure de la consanguinité, les segments d’homozygotie sont également pertinents car ils sont enrichis pour les mutations délétères. En effet, les longs segments d’homozygotie contiennent trente fois plus de mutations délétères que les segments d’homozygotie courts. Autrement dit, les mutations sont piégées dans des blocs de consanguinité (principe également connu sous le nom de « Déséquilibre de liaison » (Linkage Desequilibrium en anglais) et entraînent une augmentation de la charge génétique des mutations nuisibles au fil des générations.
Prenons par exemple l’illustration ci-dessus et imaginons que deux marqueurs génétiques sont positionnés l’un à côté de l’autre sur un chromosome. Ils seraient très probablement hérités ensemble sur le même morceau chromosomique. De même, s’ils se trouvent aux extrémités opposées du chromosome, ils sont beaucoup moins susceptibles d’être hérités sur le même morceau.
Toutes choses étant égales par ailleurs, on dit que les marqueurs proches les uns des autres sont en déséquilibre de liaison élevé entre eux et les marqueurs les plus éloignés sont en déséquilibre de liaison inférieur entre eux.
Le déséquilibre de liaison impacte la politique de sélection de l’éleveur.
En effet, l’éleveur ne peut pas sélectionner pour ou contre un seul gène étant donné que les gènes homozygotes ont tendance à se transmettre en groupes avec d’autres gènes. Les gènes sous sélection sont donc liés avec d’autre gènes dont certains peuvent être indésirables. Si un éleveur sélectionne pour (ou contre) un gène, il sélectionnera en fait tout le groupe dont fait parti le gène sélectionné y compris les gènes indésirables (tels que des gènes délétères). Il ne peut obtenir l’un sans l’autre.
La consanguinité réduit la capacité de l’éleveur à améliorer les caractéristiques de ses chiens du fait d’une diversité allélique plus faible et d’un déséquilibre de liaison plus élevé.
Les races de chiens avec une diversité génétique plus élevée ont un déséquilibre de liaison plus faible. Elles sont donc capables de répondre plus rapidement et avec plus de précision à la sélection appliquée par l’éleveur que les populations consanguines. Certes, il y a plus de variation (moins d’homogénéité), mais la possibilité de gain génétique (amélioration d’une caractéristique par la sélection) est beaucoup plus grande.
Dans la vie, vous pouvez acheter ce que vous voulez si vous avez suffisamment d’argent pour vous le permettre…
Si l’argent est la monnaie du commerce (parce qu’il permet d’acheter des choses), alors la diversité allélique est la monnaie de la sélection en élevage. Les éleveurs ne peuvent pas améliorer une caractéristique s’il n’y a pas suffisamment de variations dans les allèles pertinents entre les individus.
Consensus sur les méthodes de calcul ROH

Bien qu’il semble y avoir un accord général sur le fait que ROH est actuellement la méthode de choix pour estimer le niveau de consanguinité à partir de l’ADN, il existe encore des discussions sur la taille des blocs d’homozygotie à inclure dans le calcul (Sams & Boyko 2018). Certes, les blocs les plus longs représentent la consanguinité la plus récente et doivent être pris en compte, mais faut-il également inclure la consanguinité plus ancienne ? Cela peut représenter une différence substantielle dans les estimations de consanguinité, et pour l’instant il n’y a pas de consensus sur ce qui fournit la meilleure estimation. En attendant, des comparaisons doivent être faites entre individus ou entre races en utilisant la même taille minimale de bloc, ce qui devrait fournir des estimations comparables de la consanguinité la plus récente.
Embark
Embark utilise un chipset de 230.000 marqueurs SNP d’une résolution de 1,200 kb qui lui permet de détecter une consanguinité ancienne remontant jusqu’à 40 générations (bien mieux que n’importe quel pédigree et avec des informations fiables, contrairement à celles d’un pédigree)… La plupart du contenu d’Embark provient de la gamme de chipsets Illumina CanineHD. Ce chipset est déjà très largement utilisé dans le domaine de la recherche et de la génétique. Il a été développé par la Broad Institute (communauté multidisciplinaire de chercheurs dont la mission est d’améliorer la santé humaine). Embark y a également ajouté des marqueurs supplémentaires pour la santé et les traits.
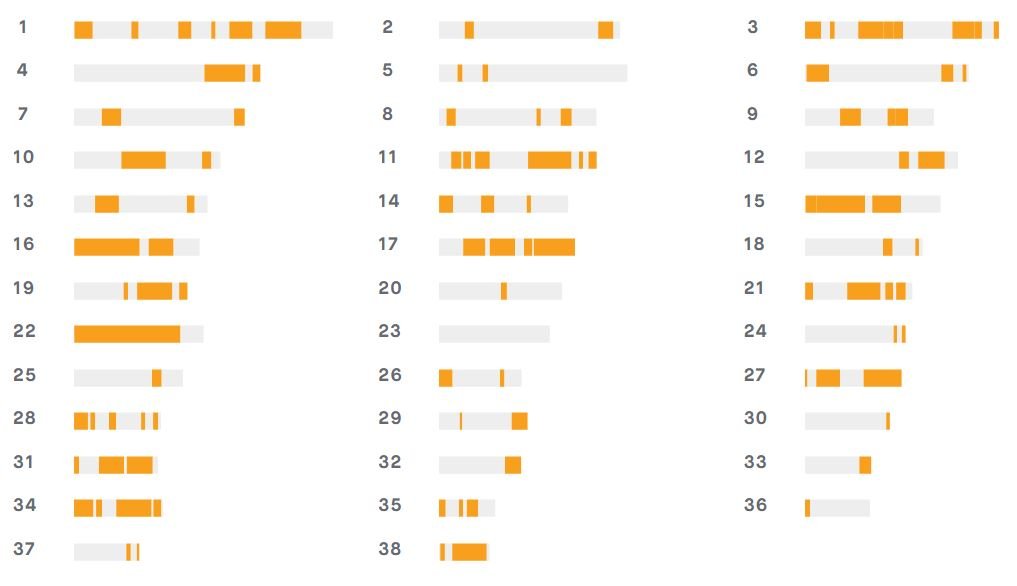
Taux de consanguinité génétique : 37%
Le “COI génétique” est excellent pour comparer l’homozygotie réelle et détecter des zones de consanguinité non visibles au pedigree. Mais ce n’est pas un diagnostic santé et il ne peut être comparé que si les marqueurs SNPs sont identiques entre les différentes plateformes, en l’absence de standardisation.
Embark est un outil puissant, mais beaucoup l’utilisent comme un tampon “qualité”. C’est exactement l’erreur : on prend un outil de mesure et on le transforme en argument marketing… Or Embark est une mesure utile d’homozygotie et un panel santé partiel, qui doit être intégré dans une stratégie.
Embark aide à objectiver l’homozygotie (ROH) et à réduire certains risques monogéniques. Mais il ne remplace ni la démographie de population, ni le pilotage de diversité, ni la parenté (kinship).
Le coefficient AVK
AVK est l’acronyme allemand pour AhnenVerlust-Koeffizient, qui signifie «Coefficient de perte d’ascendance». Ce coefficient, inventé par le Professeur Schlegel de l’Université de Vienne, en Autriche, permet d’évaluer la variabilité d’ancêtres d’après les informations du pedigree.
L’avantage de ce coefficient est sa facilité de calcul comparé au coefficient de consanguinité. De plus il n’existe pas d’options différentes de calcul contrairement au taux de consanguinité : les données sont facilement comparables !
Comment le coefficient AVK se calcule t-il ?
Il se calcule principalement sur 5 générations même si rien n’interdit de le calculer sur une profondeur de pédigree plus importante.
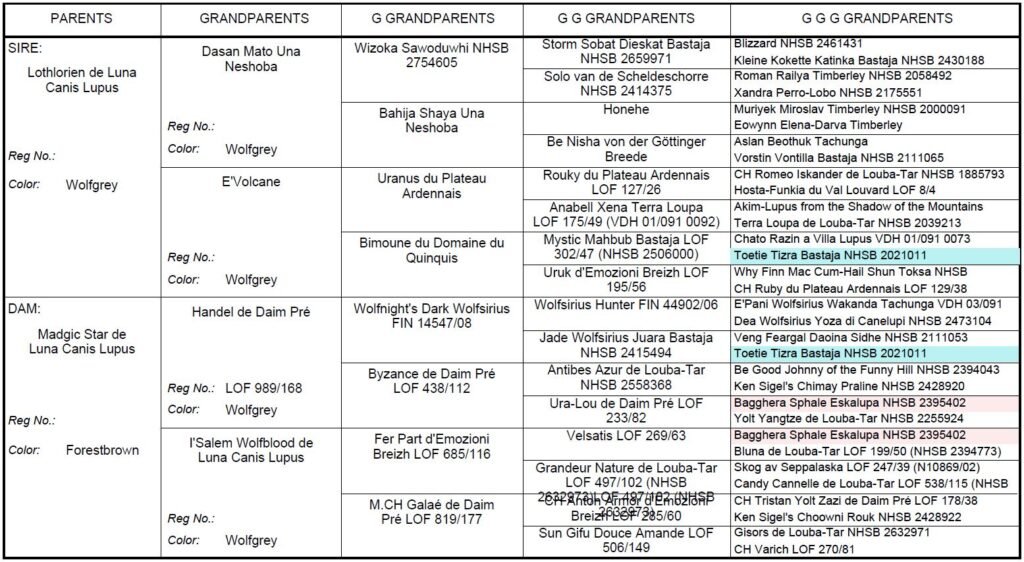
Prenons l’exemple du pédigree de cinq générations ci-dessus.
Un pédigree de cinq générations implique 62 chiens.
Deux ancêtres apparaissent deux fois (Toetie Tizra Bastaja en couleur bleue et Bagghera Sphale Eskalupa en couleur rose).
Sur les 62 chiens, il ne reste alors que 60 chiens uniques (62-2).
Les deux ancêtres sont appelés des ancêtres répétés.
Un ancêtre répété peut aussi bien apparaître uniquement côté père ou uniquement côté mère ou bien les deux. Contrairement au taux de consanguinité qui ne tient compte que des ancêtres communs (se présentant côté père ET côté mère). Le taux de consanguinité n’aurait ici tenu compte que de Toetie Tizra Bastaja, car Bagghera Sphale Eskalupa n’apparaît que côté mère.
L’AVK est un taux de variabilité ancestrale dans un pédigree.
Il permet d’apprécier la force de sélection appliquée par l’éleveur sur ses combinaisons.
L’AVK serait ici de 96.77% (60/62×100).
- Un AVK égale à 100% signifie que chaque ancêtre composant le pédigree 5 générations n’apparaît qu’une seule fois et que par conséquent, la variabilité des ancêtres est maximale.
- Un AVK inférieur à 100% est toujours une indication qu’un ou plusieurs ancêtres apparaissent plusieurs fois dans le pedigree.
- 70% est un seuil jugé comme étant une limite basse en dessous de laquelle il n’est pas recommandé de descendre. 70% correspond à 43 chiens uniques sur les 62 chiens composant un pedigree 5G.
- L’ AVK n’est pas corrélé au taux de consanguinité. Il ne s’agit pas du nombre d’ancêtres communs mais du nombre d’ancêtres uniques.
- Cependant, un mauvais AVK (inférieur à 70%) entraînera souvent un piètre résultat de consanguinité
Quand on n’a pas d’ADN et que le COI est trop court, on a besoin d’un indicateur simple pour éviter le pire : répéter les mêmes ancêtres encore et encore. L’AVK ne remplace pas la génétique, mais il nous donne une alarme utile sur la diversité d’ascendance.
L’AVK est un outil de bon sens : il aide à limiter la concentration d’ancêtres. Mais pris seul, il peut nous induire en erreur (ancêtres “différents” mais très apparentés).
Les PedCharts
Ils ont l’avantage de présenter l’intégralité du pédigree d’un chien de manière simplifiée. Malgré le nombre très important de générations impliquées, le pedchart est plus lisible que ne l’aurait été un pedigree classique. En effet, chaque ancêtre n’y est représenté qu’une seule fois (même dans le cas d’ancêtres communs apparaissant plusieurs fois dans un pédigree). Cela permet de remonter jusqu’aux fondateurs de la race et de mettre en évidence la diversité des ancêtres d’un chien. Un pédigree avec peu de diversité d’ancêtres aura tendance à avoir une forme étroite en hauteur.
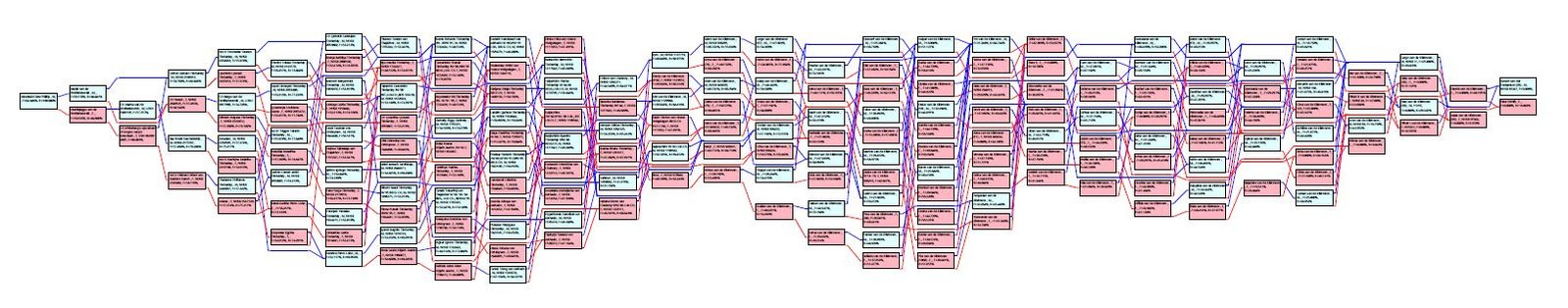

Ces PedCharts sont produits par le logiciel que j’utilise : Pedigree Publisher de Breedmate.
En bleu, les mâles et en rose, les femelles.
A gauche du Pedchart, le chien en question. Tout à droite du Pedchart, le couple fondateur de la race. Entre les deux, les différents ancêtres impliqués dans l’arbre généalogique du chien étudié.
Chaque ancêtre présente son taux de consanguinité toutes générations (F) ainsi que son coefficient de relation (COR).
L’analyse de ces deux PedCharts met en évidence les différences dans l’histoire respective de ces deux chiens-loups de Saarloos « X » et « Y ».
Si l’on observe les origines de ses deux chiens, ils ont la même histoire.
Cependant, le chien « X » est issue d’un très fort degré de consanguinité. Bien illustré par la forme étroite de la quasi totalité du pedchart.
Alors que le chien « Y » bénéficie d’une diversité d’ancêtres plus importante sur la deuxième moitié du PedChart. Par conséquent la forme du PedChart s’évase. Pour autant, ces deux pedcharts illustrent une forte consanguinité car la diversité des ancêtres est faible.
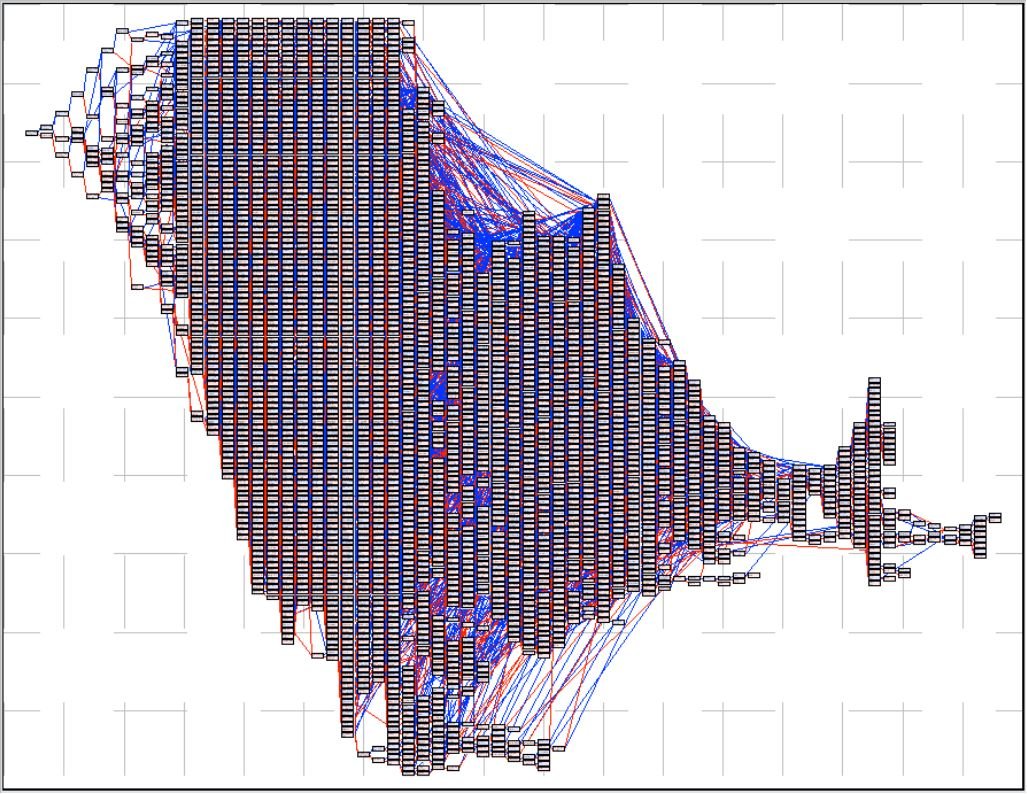
A titre de comparaison, ci-dessus le pedchart d’un Golden-Retriever ! Ce pedchart implique un nombre beaucoup plus importants d’ancêtres uniques (bien mis en évidence par la forme plus large en hauteur comparée aux deux précédents pedcharts) ainsi qu’un nombre plus important d’ancêtres fondateurs (à droite). Conséquence : un taux de consanguinité moindre pour ce Golden-Retriever comparé aux deux chiens-loups de Saarloos précédents.
Le coefficient de parenté (Kinship)
Si le but est de préserver une race, le COI ne suffit pas. On peut avoir des COI “acceptables” tout en détruisant la diversité en reproduisant les mêmes lignées. La parenté (kinship) et surtout le mean kinship (MK) servent à piloter la diversité à l’échelle de la population.
Vous envisagez de faire un mariage entre deux individus apparaissant sur un pedigree. Cependant vous ne réalisez pas leur niveau de parenté.
Sont-ils peu ou trop apparentés ?
Le coefficient de parenté (Kinship Coefficient en anglais) peut vous y aider.
Le coefficient de parenté est lié au coefficient de consanguinité. En fait, le coefficient de consanguinité d’une portée est le coefficient de parenté des parents.
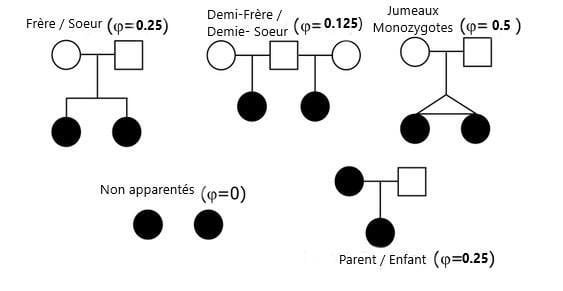
Le coefficient de parenté en génétique des populations utilise le symbole grec phi, comme sur cette figure.
Le coefficient de parenté peut être calculé à partir d’une base de données généalogiques, comme pour les coefficients de consanguinité.
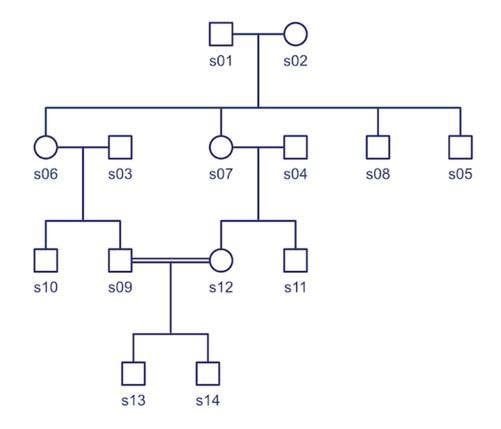
Regardons ce pedigree très simple contenant 14 chiens. Peut-être envisagiez-vous de faire un mariage entre les chiens s14 et s07, mais vous craignez qu’ils soient trop apparentés. Vous voulez savoir à quel point ces deux chiens sont étroitement liés – c’est à dire génétiquement similaires. Le coefficient de parenté vous le dira. Voici comment.
Le coefficient de parenté est toujours une comparaison de deux chiens. Vous pouvez calculer le kinship pour chaque paire de chiens du pedigree et les afficher sous forme de matrice Excel. Chaque chien est répertorié sur le côté et également sur le dessus. Le nombre dans la case où la colonne et la ligne d’une paire de chiens se croisent est le coefficient de parenté de cette paire. Les cases blanches sur la diagonale sont les chiens comparés à eux même.
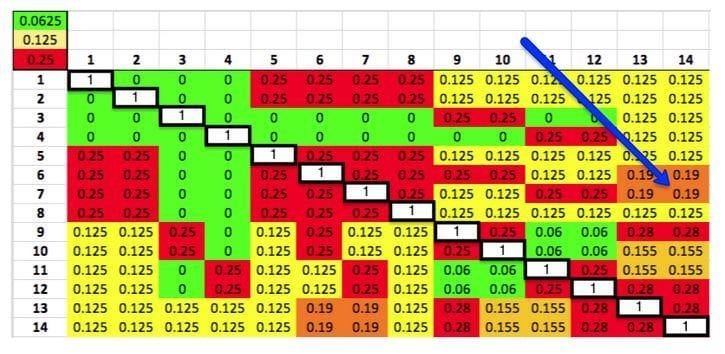
Dans le tableau ci-dessus le coefficient de parenté entre les chiens 14 et 7 est de 0,19 – soit 19%. Ainsi pour un allèle pris au hasard à un locus donné sur l’ADN des chiens 14 et 7, la probabilité de choisir deux allèles identiques hérités d’un ancêtre commun est de 0,19 – soit 19%. Ainsi, la probabilité que la progéniture issue de cette paire de chiens soit homozygote à cet allèle est de 19%. Le coefficient de consanguinité de la portée de chiots serait de 19%.
Kinship/MK nous donnent une boussole population : qui reproduire (et avec qui) pour ralentir la perte de diversité. C’est l’antidote au “meilleur mâle du moment”, le fameux mâle populaire..
Les véritables niveaux de consanguinité des chiens de race

Selon moi, l’habitude dorénavant bien ancrée de lire des taux de consanguinité calculés sur des pédigrees peu profonds occulte la vision globale du réel état du niveau de consanguinité d’une race. Et plus généralement de la situation particulière du chien de race.

Ci-dessus, un graphique synthétisant 227 races de chiens. Il est important de noter les niveaux de consanguinité représentés par les lignes horizontales de couleurs verte, jaune et rouge.
- La ligne orange représente un niveau de consanguinité de 6.25% (correspondant à un mariage entre cousins).
- En rouge représente un niveau de consanguinité de 12.5% (correspondant à un mariage entre demi-frère et demi-sœur).
- La ligne jaune représente un niveau de consanguinité de 25% (correspondant à un mariage entre frère et sœur).
- En bleu, le niveau de consanguinité du chien-loup de Saarloos.
Près de la moitiée des 227 races qui composent ce tableau ont un taux de consanguinité supérieur ou égale à 25%. C’est à dire que pour beaucoup de races de chiens, leur élevage implique un étalon et une lice qui sont au moins plus similaires génétiquement que des frères et sœurs…
Une toute petite poignée de races se situent en-dessous de 10% de consanguinité démontrant ainsi qu’une race peut préserver un type et son caractère sans utilisation excessive de consanguinité.

Les niveaux de consanguinité chez les autres animaux domestiques

Nous avons une idée plus précise du niveau de consanguinité du chien de race. Aussi il serait intéressant de savoir comment elle se situe en comparaison avec d’autres races domestiquées… Le graphique ci-dessous s’intéresse cette fois-ci aux chevaux et dans les mêmes conditions que pour celui des chiens.
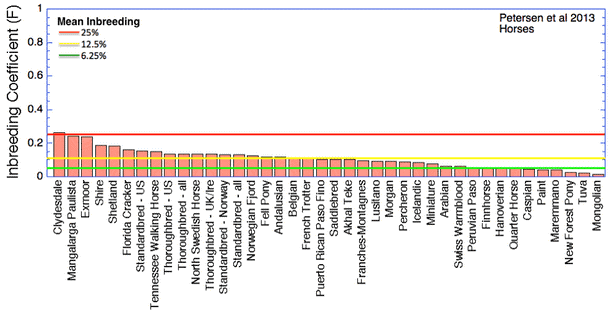
La majeure partie des races de chevaux présentes dans ce tableau se situe au plus près des 12.5% de consanguinité. Le dernier tiers se situe en-dessous des 6.25% de consanguinité… Il semble donc que la consanguinité du chien de race est beaucoup plus importante que chez d’autres animaux domestiqués. Ceci peut être expliqué par au moins deux raisons :
- Contrairement aux races de chiens, d’autres races domestiquées sont élevées avec un objectif de production. Par exemple, l’élevage de vaches en rapport à un objectif d’augmentation de production de lait. Pour atteindre cet objectif, un programme d’élevage est mis en place qui va mélanger les races. Une fois l’objectif atteint, le programme d’élevage est arrêté. Puis commence alors un programme de multiplication de ces vaches « améliorées » pour maintenir les performances de production de lait.
Alors que chez le chien de race, le terme « élevage » implique de ne pas mélanger les races. Les livres des origines sont fermés dès la création de la race. Tout se fait par la suite en consanguinité. - Cependant, la sélection appliquée par les éleveurs va par la suite expliquer les différences de niveau de consanguinité du chien de race et celui du cheval par exemple. Nous développerons plus loin dans cet article, cet aspect capital.
L’hypertype
Parmi les mythes et légendes qui pullulent dans le monde cynophile, il y en a un qui a la vie dure et qui consiste à faire croire que la consanguinité est nécessaire pour préserver le type…
Or, le type a été fixé dans les premières générations qui ont suivi la création de la race au travers de sa population fondatrice. Son pool génétique d’origine définit les qualités physiques et comportementales de la race mais également son stock de diversité génétique de départ. Une consanguinité supplémentaire augmente seulement le niveau d’homozygotie dans les autres gènes du chien. Il y a de très nombreuses mutations récessives qui se cachent parmi les milliers de gènes dans un chien. Pour des questions bassement esthétiques ou comportementales, l’excès de consanguinité produira par inadvertance des mutations par paires (homozygotie) pour certains gènes. Parmi ces gènes devenus homozygotes, certains seront bénéfiques (la plupart du temps, ce sont les gènes « visibles », ceux qui impactent le phénotype) mais certains seront délétères (et la plupart du temps, ils ne sont pas visibles immédiatement et produiront des maladies qui peuvent être monogéniques ou polygéniques).
L’excès de consanguinité n’est pas nécessaire pour préserver un type et une race peut avoir un phénotype très reconnaissable avec une consanguinité inférieure à 10%
Qu’y a-t’il de plus à fixer qui n’est déjà mentionné dans le standard ?
Quelle est cette quête de types dans le type ?
Les dérives de sélections sont très souvent à l’origine des hypertypes apparus dans certaines races de chiens (Berger Allemands, races brachycéphales etc.). Leurs conséquences sur la santé sont désastreuses…





La course à l’hypertype fait partie des grands maux de l’élevage canin, dont les seuls objectifs sont la recherche du spectaculaire, de l’original et la réussite aux concours de beauté, au détriment de la santé et du bien-être des animaux.
Je me demande bien pourquoi, en tant qu’éleveurs, exposants ou juges, nous avons souvent du mal à admettre que nous nous habituons aux changements qui interviennent au sein des races. Dans certains cas, ces changements sont tellement incrustés dans une race qu’ils sont élevés au rang de caractères désirés alors qu’ils n’apportent absolument rien ni au type ni à la fonction. En général ces changements commencent avec des chiens présentés par des exposants ou des présentateurs célèbres qui posent des jalons que tout le monde suivra allègrement. Comment les changements s’opèrent-ils ? Ils débutent chez l’éleveur qui élève une portée jusqu’à ce que les chiots soient en âge d’être évalués. L’éleveur qui connaît parfaitement la race examine chaque chiot en gardant à l’esprit le standard et ce qui est correct pour la race. Pour la plupart des races, la » modération » est de mise à plus d’un titre. Considérer tous les caractères d’un quelconque animal avec modération contribuera à l’équilibre de cet animal et au sentiment que » tout colle « . Toutefois, il peut advenir qu’un chiot ait quelque chose de particulier qui attire l’œil et, inévitablement ce « quelque chose» est l’exagération d’un caractère… un cou trop long, une tête trop fine, une angulation des postérieurs excessive… et c’est là que se situe le danger. L’éleveur qui connaît parfaitement la race pour y être profondément impliqué considèrera ce » quelque chose » pour ce qu’elle est, c’est à dire une exagération et écartera ce chiot qui ne correspond pas au type. Cependant, beaucoup d’autres penseront que cette exagération qui attire inévitablement leur œil ne manquera pas d’attirer celui du juge dès que le chien entrera sur le ring. Et c’est ainsi que ce chiot est gardé, choyé, éduqué et toiletté. Aussitôt qu’un chien d’un type «déviant» commence à gagner, la machine est lancée. Ce chien fait son entrée sur le ring d’honneur et d’autres juges font remarquer sa magnifique ossature, son long cou, ses fabuleuses angulations, son corps ultra court, son port de queue quand bien même ces caractères ne sont pas spécifiques de la race. Ils le récompensent quand ils le rencontrent et les autres lui emboitent allègrement le pas. Les éleveurs débutants voyant ce chien ainsi que les prix qu’il remporte pensent qu’ils doivent le prendre comme exemple. Ils se précipitent pour utiliser ce chien et en quelques années, le type « déviant » a la main mise sur la race. En très peu de temps, un juge peut se trouver devant six chiens dont cinq du «nouveau» type et un absolument correct . Un juge bien informé et au fait de la race pourra affirmer « celui-ci est correct, les cinq autres ne le sont pas ». Par contre beaucoup d’autres juges connaissant peut-être moins la race, choisissant la solution de facilité penseront que les cinq doivent être corrects puisqu’ils sont majoritaires et le sixième rentre chez lui sans récompense. A ce moment-là, l’éleveur dévoué qui s’est appliqué à maintenir le type et améliorer la qualité commence à se sentir frustré et cela se comprend. Il sait que ce qu’il produit est correct mais le nombre de ceux qui s’éloignent du standard est tellement grand que les juges et les exposants suivent la mode alors que lui, reste seul à nager à contre courant. Cela s’est produit pour différentes races en Grande Bretagne et ailleurs. J’ai vu beaucoup d’éleveurs « des temps anciens » exposer de moins en moins. Il leur semblait tout simplement qu’il ne servait à rien de présenter leurs chiens à des juges qui ne connaissent pas les caractères essentiels de la race. C’est pourtant ces éleveurs-là qui DEVRAIENT exposer massivement afin que le public « vierge » puisse voir et apprécier ces chiens. Lorsque des chiens avec des défauts ( défauts qui attirent l’œil ) continuent à gagner et reproduire, les nouveaux éleveurs n’ont aucune raison de corriger ou améliorer les spécimens. Pourquoi le feraient-ils ? Ces chiens gagnent. Les propriétaires des chiens « à la mode » ont en général la parole et avancent des arguments convaincants sur l’évolution naturelle ainsi que les « améliorations » évidentes. Parfois des esprits obstinés vont jusqu’à vouloir influencer le club de race, demandant une révision du standard afin que celui-ci s’adapte à la « nouvelle mode » … un véritable crime. Sans oublier bien sûr la puissance des annonces ! Malheureusement les nombreux changements auxquels nous avons assisté ont les faveurs du public non initié. Qu’est-ce que ça peut faire si un chien est trop poilu, trop droit, trop court, trop long ? Il paraît joli et les juges aiment cette apparence ! Bien que de nos jours le but des expositions soit davantage une course aux récompenses plutôt qu’une préservation de la race, le ring demeure la vitrine des éleveurs. Il serait bien triste de penser que les véritables éleveurs qui préservent le vrai type de la race avec enthousiasme ne puissent trouver des chiens capables de transmettre leur beauté à la génération suivante. »
Andrew H. Brace, journaliste canin et juge anglais.
L’hypertype a également une conséquence sur le génotype. En effet, les éleveurs addictes des expositions, ont historiquement peu pris en compte la manière dont la sélection de la prochaine portée affecte la génétique de la race, ce qui met effectivement les objectifs à court terme en contradiction avec la durabilité à long terme de la population reproductrice. C’est cette focalisation sur les programmes d’élevage individuels qui a poussé la génétique de nombreuses races dans une direction qui, en fin de compte, rend la sélection de plus en plus difficile et l’élevage de moins en moins durable.
La plupart des discours sur la consanguinité évitent le sujet qui fâche : ce ne sont pas les accouplements “cousins” qui tuent une race, c’est la dynamique démographique et la sélections extrême. Mâles populaires et hypertypes sont deux manières différentes d’acheter du court terme et de vendre le long terme.
Si on ne contrôle pas la concentration de reproducteurs et la dérive esthétique, on peut afficher des COI “raisonnables” tout en détruisant la diversité et la fonctionnalité.
La consanguinité du chien de race : la roulette Russe ?

Beaucoup cherchent “la maladie” qui prouverait que la consanguinité est dangereuse. Mauvaise approche. La dépression de consanguinité est souvent une baisse globale de robustesse, difficile à attribuer à une cause unique… jusqu’au moment où les indicateurs (fertilité, mortalité, longévité) se dégradent franchement.
S’il y a des mutations parmi les milliers de gènes chez le chien de race (et vous pouvez être sûr qu’il y en a !), la consanguinité produira par inadvertance des mutations par paires (homozygotes) pour certains gènes. Ainsi ces mutations impacteront le fonctionnement d’un gène normal. Peut-être que ce gène est lié simplement à un trait physique ou bien lié au bon fonctionnement du métabolisme du chien. Pour autant, c’est la roulette russe… Le côté apprenti sorcier de la sélection appliquée par l’éleveur… D’ailleurs, aucun d’entre eux ne peut se targuer de connaître à l’avance le fruit de ses combinaisons sur le plan génétique. Et encore moins de clamer haut et fort qu’il sait ce qu’il y a dans ses lignées…
Il n’y a aucun moyen de savoir pour un éleveur si le fruit de sa sélection entraînera un effet positif (un trait physique amélioré) ou un effet négatif (mutation génétique récessive révélée par homozygotie).
D’autres animaux domestiques ont des races pourtant clairement reconnaissables. D’ailleurs leur niveau de consanguinité est beaucoup plus faible que celui du chien de race… Par contre, en ne choisissant que les “meilleurs” individus pour améliorer le phénotype, une partie de la variabilité génétique se perd. Ainsi chaque gène perdu entraîne potentiellement un effet et des dizaines de générations plus tard, la variabilité génétique du début s’est amoindrit. C’est ainsi que certaines choses ne fonctionnent plus comme elles le devraient niveau santé. Il ne s’agit pas ici de “mutations” génétique mais simplement d’un dysfonctionnement du gène “normal”. Celui-ci aurait dû faire son travail s’il n’avait pas été modifié par la sélection appliquée par l’éleveur.
Nous avons commencé avec des chiens en bonne santé puis nous avons jeté des gènes, et maintenant nous avons des chiens avec des maladies héréditaires.
Nous ne résoudrons JAMAIS ce problème en continuant de perdre davantage de gènes tout en essayant de « se débarrasser » des mutations. Le problème n’est pas que nous avons une mutation. Le problème est que nous n’avons plus de copie du gène normal. Nous devons remettre les gènes perdus dans le pool génétique afin que les choses fonctionnent comme elles le devraient naturellement.
Comment les populations d’animaux sauvages (au phénotype pourtant reconnaissable) parviennent-elles à rester en bonne santé pendant des milliers de générations ? Et sans test de santé qui plus est !!! Parce qu’ils ne perdent pas de gènes importants dans leur pool génétique. Les animaux se déplacent parfois de troupeau en troupeau. Ce qui restaure les gènes perdus d’une population qui s’était dissociée des autres.
Si une population devient isolée et que de nouveaux individus sont empêchés de s’y joindre, le pool génétique se réduit, la santé de la population se dégrade jusqu’au moment inéluctable où la population disparaît…
On peut passer des années à “ne rien voir” et quand ça casse, il est tard : diversité perdue, goulots fixés, options réduites. Le seul antidote fiable est démographique et stratégique, pas narratif.
Quelle est la valeur de consanguinité à ne pas dépasser pour le chien de race ?

Les effets négatifs de la consanguinité commencent à devenir évidents avec un coefficient de consanguinité d’environ 5%. À un niveau de 10%, il y a une perte de vitalité chez la progéniture ainsi qu’une augmentation de l’expression des mutations récessives délétères. Des niveaux de consanguinité supérieurs à 10% auront des effets non seulement sur la qualité de la progéniture, mais également des effets néfastes sur la race. Pour rappel, 6.25% correspond à un mariage entre cousins…
Pour autant, devrait-on établir une règle qui interdirait toute reproduction dont le résultat de consanguinité serait supérieur à 10 % ? Après tout, ce serait une règle logique… Malheureusement, les choses ne sont pas si simples…
Avec les outils dont nous disposons actuellement, la consanguinité ne peut qu’être estimée. À partir d’une base de données généalogiques, nous ne pouvons obtenir qu’une estimation moyenne de la consanguinité de la portée et non de la consanguinité de chaque individu au sein de la portée.
En effet, un coefficient de consanguinité généalogique est basé sur la filiation des ancêtres. Bien que tous les chiots d’une portée ont les mêmes parents, chacun aura eu une distribution d’allèles différente. Par conséquent, la diversité génétique au sein d’une portée est différente d’un individu à un autre. Le coefficient de consanguinité toute génération est une estimation de la consanguinité moyenne attendue pour une portée. Cependant la consanguinité réelle pour chaque individu peut être supérieure ou inférieure à cette moyenne. Pour cette seule raison, il serait ridicule de déclarer 10% comme seuil absolu pour les reproductions. Pour connaître le niveau réel d’hétérozygotie de chaque chiot d’une portée, il suffit de les tester sur Embark. La comparaison de leurs taux individuels de consanguinité apportera un critère supplémentaire à l’éleveur pour sa sélection d’élevage.
Mais il faut se méfier de ne pas réduire un ensemble complexe de considérations génétiques à une règle simple. En effet il y a beaucoup plus à considérer. Ainsi plus on apprend, plus on réalise à quel point il est dangereux de prendre des décisions compliquées basées sur une déclaration concise…
Outcross (ou croisement)
Source : Institute of Canine Biology
Les pools génétiques fermés imposés par les instances cynophiles pour les chiens de race pure entraînent nécessairement une consanguination, et chez de nombreuses races, le niveau moyen de consanguination est déjà élevé. C’est la raison pour laquelle l’apparition de troubles génétiques chez les chiens de race pure augmente régulièrement en même temps qu’une plus faible fertilité, des portées plus petites et une mortalité plus élevée des chiots rendent la reproduction encore plus difficile.
Votre première option est de faire le meilleur usage possible de la diversité génétique qui existe encore chez votre race. Identifiez les lignées qui ne sont pas étroitement liées aux vôtres, et même si ces animaux ne seraient pas votre premier choix en termes de type, un croisement produisant un COI plus faible sera bénéfique pour la prochaine génération en termes de santé. Une analyse génétique de la base de données de pedigrees de votre race peut vous aider à trouver ces animaux moins apparentés en utilisant ce qu’on appelle l’analyse de grappes (dendrogramme).
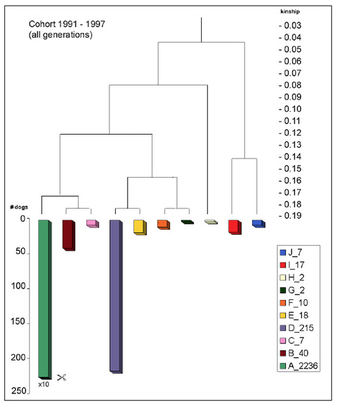
Ne présumez pas que les animaux de différentes lignées ou même de différents pays sont moins liés. Calculez le coefficient de consanguinité d’un accouplement potentiel à partir d’une bonne base de données de pedigree qui remonte aux fondateurs. Un croisement fait avec un chien qui est plus lié que vous ne le réalisez est susceptible de produire une portée avec beaucoup de mauvaises surprises.
Que se passe-t-il si votre race est si consanguine qu’il n’y a nulle part où aller pour trouver des animaux moins apparentés ? Malheureusement, de nombreuses races sont confrontées à ce problème. La diversité génétique est inévitablement perdue d’une race à chaque génération, et pour restaurer la diversité et réduire la consanguinité, vous devez remettre les gènes perdus dans le pool génétique par un chien non apparenté, probablement d’une race différente. Si votre race est déjà très consanguine et aux prises avec des problèmes de santé importants, ce n’est pas une chose triviale à faire. Les animaux à croiser doivent être sélectionnés avec beaucoup de soin. Par exemple, l’élevage d’un autre chien très consanguin, même d’une race différente, produira une progéniture qui ont tous les mêmes allèles pour les gènes qui étaient homozygotes chez le parent. La clé de la gestion des mutations récessives dans toute population est de les garder rares, de sorte que l’ajout d’animaux à la population qui partagent un grand nombre des mêmes mutations ne pose des problèmes à l’avenir. En outre, l’incorporation d’un nouveau matériel génétique dans la race nécessitera une stratégie bien conçue pour au moins les 4 ou 5 prochaines générations. Un seul croisement suivi d’un backcrossing séquentiel dans la race éliminera la majeure partie de la diversité génétique que vous espériez introduire. Vous devez certainement commencer par un plan soigneusement conçu par des généticiens avec les outils nécessaires pour le faire correctement.
Éviter des niveaux élevés de consanguinité en premier lieu est beaucoup plus facile que d’essayer de réparer les choses après que la consanguinité soit devenue un problème. Les éleveurs devraient travailler ensemble pour surveiller la consanguinité de leur race afin qu’ils puissent tous bénéficier de chiots en meilleure santé qui atteignent leurs objectifs en tant qu’éleveurs maintenant et à l’avenir.
L’outcross n’est pas une “solution miracle”, c’est un outil coûteux qui exige une stratégie multi-générations et une discipline de données. Sans ça, on ne “sauve” rien : on fabrique juste des chiens difficiles à intégrer et on perd la confiance.
Un outcross bien fait restaure des options génétiques. Un outcross mal fait détruit le type, ajoute des problèmes, et échoue politiquement. La différence : objectifs clairs, métriques, gouvernance, transparence.
Dois-je quand même me préoccuper du coefficient de consanguinité si je fais les tests de santé disponibles pour ma race ?
Définitivement oui ! Les tests de santé ne vous renseignent que sur un gène particulier pour un risque connu.
Nous pouvons savoir si un chien possède une mutation délétère seulement si nous avons un test pour cela.
La plupart des chiens de races ont de nombreuses mutations récessives cachées dans leur génome. Or la plupart ne peuvent pas être dépistées puisqu’il n’existe pas encore de tests de santé. Aucun moyen de les connaître à l’avance sauf lorsqu’elles s’exprimeront par homozygotie…
Vous élevez peut-être des chiens « testés pour la santé », mais vous produisez toujours des chiots présentant un risque important de maladies génétiques dues à des mutations récessives !
Elever des animaux en bonne santé, c’est s’inquiéter de TOUS les risques potentiels, et la seule chose dont nous pouvons être sûrs, c’est qu’il y a beaucoup plus de mutations récessives que celles pour lesquelles nous avons des tests de santé. Pourquoi investir dans les tests de santé disponibles pour votre race, puis produire une portée dans laquelle 15 %, 25 % ou 40 % des autres mutations de chaque animal seront exprimées ?
Les tests de santé et la recherche ne résoudront pas le problème tant que les éleveurs, d’une manière générale alimentent eux-mêmes ce cycle par des choix de mariages n’agissant pas sur la dépression de consanguinité.
Puis-je utiliser le coefficient de consanguination pour réduire le risque de troubles génétiques chez mes chiots ?
Est-ce que le coefficient de consanguinité peut être utilisé pour réduire le risque de troubles génétiques chez les chiots ? Absolument ! C’est exactement ce pour quoi il a été conçu. Rappelez-vous simplement que le COI est une estimation de la perte prévue de vigueur et de santé générale à attendre en raison de l’expression de mutations récessives. Sauf pendant le développement d’une nouvelle race (où les éleveurs vont vouloir utiliser la consanguinité pour fixer le type) vous devriez vous efforcer de maintenir la consanguinité en dessous de 10 % pour obtenir un bénéfice modeste avec un risque modeste.
Conduite d’élevage à adopter

- Utiliser tous les tests de santé disponibles pour éliminer tous les risques de mutations récessives dont nous connaissons l’existence.
- Puis utiliser le taux de consanguinité pour s’assurer d’avoir réduit autant que possible le risque de mutations récessives inconnues qui se cachent dans le génome de chaque chien.
- De préférence, utiliser des taux de consanguinité génétique (aujourd’hui possible avec Embark) et demander à Embark de vous calculer les taux de consanguinité génétique de plusieurs combinaisons afin que vous puissiez faire votre choix en toute connaissance des différents niveaux de risques produits par chaque combinaison.
- S’assurer à minima que les chiots produits auront un taux de consanguinité inférieur à celui de leurs parents.
Le coefficient de parenté (kinship coefficient) et le coefficient de perte ancestrale (AVK) sont d’autres indicateurs intéressants si le taux de consanguinité génétique n’est pas accessible. Il s’agira alors de valeurs prédictives de la portée et non des valeurs individuelles des chiots de la portée.
La plupart des erreurs d’élevage viennent d’un problème simple : décisions prises au feeling, puis justifiées après coup. Cette checklist nous force à décider avant, sur des critères explicites, et à documenter nos compromis. C’est la différence entre “produire une portée” et “piloter une lignée”.
Une bonne conduite d’élevage est une procédure, pas une opinion : filtres, métriques, arbitrage, transparence, suivi. Si on ne peut pas expliquer notre choix en 10 lignes factuelles, c’est qu’on ne l’a pas vraiment décidé.
La mauvaise utilisation de la consanguinité dans les pratiques d’élevage
(Source : Institute of Canine Biology, Carole Beuchat PhD)
Pour produire des chiens en bonne santé, nous devons réduire le plus possible le risque de problèmes causés par TOUTES les mutations récessives. Celles que nous connaissons et celles que nous ignorons.
Vous devez vous rappeler que le coefficient de consanguinité n’est pas une mesure du niveau de santé. C’est une mesure du RISQUE. Avec ou sans tests de santé, c’est la meilleure façon de juger du niveau de risque génétique que vous prenez lorsque vous produisez une portée.
Un peu d’histoire
Grâce à une série d’expériences, Johann Gregor Mendel, botaniste Autrichien à la fin du 19ème siècle, a définit la manière dont les gènes se transmettent de génération en génération (lois de Mendel) ainsi que leur implication dans l’hérédité des traits chez les plantes et les animaux. Les éleveurs ont pu ainsi faire de simples expériences de sélection pour révéler si les gènes pour des traits spécifiques étaient dominants ou récessifs. Ce qui leur a permis de pratiquer des sélections produisant des traits prévisibles.
Bien que cela ait éliminé une partie du mystère de la sélection pour de nombreux traits, pour d’autres, leur prédiction est restée difficile à atteindre. Les éleveurs ont réalisé que, dans certains cas, la génétique d’un trait devait être plus complexe.
En effet, certains traits sont déterminés par le cumul de plusieurs gènes et non par un seul (traits polygéniques). C’est un généticien universitaire nommé Jay Laurence Lush qui dans les années 1930, s’est rendu compte que, pour de nombreux traits, il pouvait y avoir beaucoup de variations d’un individu à l’autre. Autrement dit, certains traits n’étaient pas « binaires ».
Par exemple, il ne peut y avoir deux tailles possibles pour un animal déterminé. En effet entre le plus petit individu et le plus grand, il y a toute une variation de différentes tailles.
Ainsi il s’est rendu compte que de nombreux traits étaient polygéniques. Aussi, il a commencé à réfléchir à la manière dont la génétique pouvait expliquer cela.
Les traits polygéniques
Jay Laurence Lush découvre qu’une variation génétique fournit la matière première pour la sélection. De plus une partie de cette variation est « cumulative« . Ainsi un grand chien accouplé à une grande chienne pourrait produire une progéniture plus grande que l’un ou l’autre de ses parents.
L’observation de Lush a lancé un nouveau concept dans la génétique moderne, appelé « génétique quantitative ». Ce concept est appliqué spécifiquement aux traits qui varient continuellement. Les progrès dans la compréhension de la génétique quantitative ont par la suite révolutionné l’élevage d’animaux. Ils ont été à la base des améliorations spectaculaires de la reproduction animale. Elle est devenue le système moderne d’élevage commercial du bétail et d’autres animaux.
Bien sûr, les chiens de race présentent de nombreux exemples de traits qui ont été façonnés par une reproduction sélective et qui tire parti de la variation génétique cumulative. Ainsi la taille d’un animal en est un exemple. A ce sujet, les chercheurs ont identifié plusieurs gènes pouvant être associés à une variation de la taille corporelle chez le chien. Mais des dizaines voire des centaines d’autres gènes ayant de minuscules effets individuellement (modèle polygénique infinitésimal) ont en fait, une influence importante collectivement.
Un exemple : comment améliorer la rapidité de course ?
Prenons pour exemple une race de chien dont la rapidité de course doit être améliorée.
Comment les éleveurs pourraient-ils sélectionner pour élever des chiens toujours plus rapides ?
Une option serait de marier uniquement le meilleur mâle avec la meilleure femelle.
Est-ce qu’une partie de la progéniture sera plus rapide que l’un ou l’autre des parents ? Peut-être.
Certains pourraient-ils être aussi plus lents ? Peut-être également…
Alors l’éleveur choisit d’accoupler le mâle le plus rapide de la portée avec la femelle la plus rapide. Il sélectionne à nouveau les plus rapides de leur progéniture. L’éleveur pourrait ainsi obtenir des chiens plus rapides que les deux premiers parents. Cependant au fil des générations, de consanguinité et de sélections, les progrès commenceront à se stabiliser. C’est l’élevage basé uniquement sur le concept du « meilleur des meilleurs ».
Malheureusement, une grande partie de la diversité génétique d’origine des ancêtres de la première génération aura été perdue. En effet, les chiots de chaque portée partageront une part de plus en plus grande des mêmes gènes à chaque génération. L’éleveur ne pourra plus obtenir de chiens de plus en plus rapides. Pourquoi ? Par les effets délétères de la consanguinité, l’éleveur a éliminé la variabilité génétique nécessaire pour produire une amélioration d’un trait continu en délaissant la variabilité génétique cumulative.
L’éleveur se retrouve finalement dans une impasse génétique faute de ne s’être pas constitué de « stock » minimum d’individus présentant une variabilité génétique suffisante.
La sélection de groupe plutôt que la sélection individuelle
Une sélection de groupe et non individuelle augmentera la variabilité génétique de ces chiens. Ainsi l’héritage aléatoire de cette variation chez la progéniture poussera les performances de course. En effet, pour améliorer un trait polygénique, une variabilité génétique est nécessaire pour la sélection. Au lieu de sélectionner un seul meilleur chien, une sélection d’un groupe de chiens tous plus rapides que la moyenne est préférable.
Est-il possible de produire des chiens de plus en plus rapides pour toujours ?
Probablement pas. Même avec une variabilité génétique préservée, les limites de la conception et de la physiologie s’imposeront à un moment donné. Mais ce type d’élevage sélectif permettra sans aucun doute d’améliorer la plupart des traits polygéniques chez les chiens.
Quels sont les traits recherchés qui sont polygéniques ?
La taille, le comportement, le tempérament, la capacité au travail, la qualité du pelage, la couleur des yeux, la longueur d’oreille, la longueur du museau, les angulations etc. La liste est longue.
Ainsi nous arrivons au problème sous-jacent de la consanguinité : la surexploitation des reproducteurs dit populaires dans la reproduction…
Les reproducteurs populaires
Lorsque les éleveurs ne sélectionnent uniquement que le «meilleur» individu à reproduire, ils sélectionnent la combinaison de gènes et d’environnement qui a produit ce qu’ils perçoivent comme étant le « meilleur ».
Cependant, d’autres chiens pourraient également présenter un intérêt pour le trait recherché. Mais malheureusement, ils ne sont pas le « meilleur ». Ils ne sont donc pas sélectionnés pour contribuer à l’amélioration de ce trait… Les gènes de ces chiens seront perdus si l’éleveur ne sélectionne que le « meilleur » chien.
Au lieu de continuer dans cette culture du « meilleur » chien, il serait mieux d’élever un groupe de chiens qui seraient les plus performants sur le critère recherché. Ainsi, la variabilité génétique est préservée tout en continuant à améliorer les différents traits recherchés.
Redéfinir la notion de « meilleur » est primordial. Sélectionner les meilleurs individus à reproduire mais en termes de génétique de groupe.
Vous avez sans aucun doute remarqué que cette méthode d’élevage, où le « meilleur » est défini comme un groupe d’individus au lieu d’un seul chien, n’est pas la façon dont les éleveurs de chiens élèvent habituellement. En effet ils choisissent généralement de garder et d’élever uniquement LE chien qu’ils perçoivent comme étant le « meilleur » en ignorant les effets possibles de l’environnement sur leur évaluation des autres chiens apparemment inférieurs.
Ainsi ils rétrécissent le patrimoine génétique et augmentent la consanguinité à chaque génération. Leur capacité de continuer à améliorer les traits de leur cheptel s’en trouvera diminué à cause du déséquilibre de liaison. Les conséquences impactent la santé et pas uniquement les traits physiques et comportementaux.
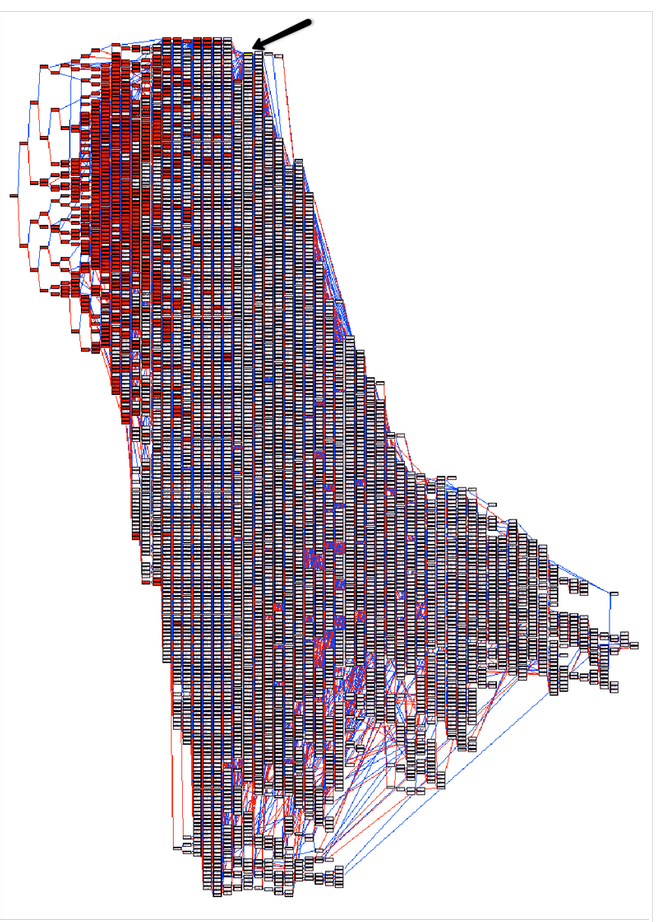
Ci-dessus, un bel exemple de l’impact catastrophique des étalons populaires. Il aura suffit d’un seul mâle populaire signalé par la flèche noire pour rétrécir le patrimoine génétique des dernières générations de ce PedChart. Tous les descendants en rouge sont les descendants directs de ce mâle multi-champion des concours de beauté à la mode…
Conclusion

« La majorité des éleveurs pensent agir en faveur de la conservation, à partir du moment où ils ne croisent que des chiens de race pure. Quant à l’amélioration, la plupart des éleveurs pensent aussi qu’ils peuvent y arriver, en laissant les «meilleurs » chiens contribuer le plus possible à la reproduction des générations futures. Ils pensent aussi que cette approche permet d’éradiquer les problèmes de santé et de bien-être chez les chiens de race.
La réalité de l’élevage est cependant toute différente. Le fait est qu’en dépit de tous nos efforts, le pourcentage d’animaux souffrant de troubles héréditaires ne cesse d’augmenter, et ne semble pas diminuer.
Toutes nos tentatives pour améliorer la santé et le bien-être de la population des chiens de race via cette sélection, aboutit quasiment au néant. »
Lr. Ed. J. Gubbels, Généticien
Ces propos alarmants du Dr. J. Gubbels (généticien), ont été tenus lors d’une conférence qu’il a animé au cours de la Mondiale d’Amsterdam de 2002.
Dans une race à livre fermé, la consanguinité n’est pas une “option”, c’est un continuum. On choisis soit de l’ignorer (et d’accélérer la perte), soit de la mesurer, de ralentir la perte de diversité, et de décider avec une stratégie claire.
Si on ne devait retenir qu’une chose : un taux de consanguinité isolé ne suffit pas. Une bonne décision d’élevage, c’est un compromis explicite entre santé, diversité, et objectifs — documenté, assumé, et cohérent sur plusieurs générations.
Merci pour cette pépite !! Votre article est très complet, didactique et quel travail ! Sincèrement MERCI !!
Anonyme = Isabelle, présidente de l’association « Passion Spitz Loup » 🙂
Merci pour vos encouragements Isabelle ^^
Merci de ce magnifique article très bien documenté qui ne fait que confirmer mon expérience et mes écrits http://hogandesvents.nutritionverte.com/articles.html
Bien cordialement
Mathieu Mauriès